
Western Michigan University Western Michigan University
ScholarWorks at WMU ScholarWorks at WMU
Masters Theses Graduate College
4-1975
The Effect of Audible Sound on the Germination and Root The Effect of Audible Sound on the Germination and Root
Elongation of Selected Seedlings Elongation of Selected Seedlings
Micheal Dennis Walton
Western Michigan University
Follow this and additional works at: https://scholarworks.wmich.edu/masters_theses
Part of the Biology Commons
Recommended Citation Recommended Citation
Walton, Micheal Dennis, "The Effect of Audible Sound on the Germination and Root Elongation of Selected
Seedlings" (1975).
Masters Theses
. 2528.
https://scholarworks.wmich.edu/masters_theses/2528
This Masters Thesis-Open Access is brought to you for
free and open access by the Graduate College at
ScholarWorks at WMU. It has been accepted for inclusion
in Masters Theses by an authorized administrator of
ScholarWorks at WMU. For more information, please
contact [email protected].

THE EFrriCT OF AUDIBLE SOUND ON THE GERMINATION AND
ROOT ELONGATION OF SELECTED SEEDLINGS
by
Micheal Dermis Walton
A Thesis
Submitted to the
Faculty of The Graduate College
in partial fulfillment
of the
Degree of Master of Arts
Western Michigan University
Kalamazoo, Michigan
April 1975
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

ACKNOWLEDGEMENTS
I' am very grateful for the advice and assistance I received
in the undertaking of this project to Dr. Leo C. Vander 3eek and
the other members of my thesis committee, Dr. Richard Y,\ Pippen
and Dr. Eugene 3emstein.
A- special thanks to a good friend iir. F e m e (3ud) Ellis
without whose help this task would have been much more difficult.
His assistance in the setting up of the experiment and in running
the data through the computer was of immeasurable value.
Finally, a word of thanks to Mr. Parrish, who works in the
electronics laboratory, for his help in repairing and replacing
some of the sound equipment which was necessary before the experiment
could be started.
Micheal Dennis Walton
mi
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

IN FO R M ATIO N TO USERS
This material was produced from a microfilm copy of the original document. While
the most advanced technological means to photograph and reproduce this document
have been used, the quality is heavily dependent upon the quality of the original
submitted.
The following explanation of techniques is provided to help you understand
markings or patterns which may appear on this reproduction.
1.T h e sign or "target" for pages apparently lacking from the document
photographed is "Missing Page(s)". If it was possible to obtain the missing
page(s) or section, they are spliced into the film along with adjacent pages.
This may have necessitated cutting thru an image and duplicating adjacent
pages to insure you complete continuity.
2. When an image on the film is obliterated with a large round black mark, it
is an indication that the photographer suspected that the copy may have
moved during exposure and thus cause a blurred image. You will find a
good image o f the page in the adjacent frame.
3. When a map, drawing or chart, etc., was part of the material being
photographed the photographer, followed a definite method in
"sectioning" the material. It is customary to begin photoing at the upper
left hand corner of a large sheet and to continue photoing from left to
right in equal sections with a small overlap. If necessary, sectioning is
continued again — beginning below the first row and continuing on until
complete.
4. The majority of users indicate that the textual content is of greatest value,
however, a somewhat higher quality reproduction could be made from
"photographs" if essential to the understanding of the dissertation. Silver
prints of "photographs" may be ordered at additional charge by writing
the Order Department, giving the catalog number, title , author and
specific pages you wish reproduced.
5. PLEASE NOTE: Some pages may have indistinct print. Filmed as
received.
Xerox University Microfilms
300 North Zeeb Road
Ann Arbor, Michigan 48106
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

MASTERS THESIS
M-7033
WALTON, Micheal Dennis
THE EFFECT OF AUDIBLE SOUND ON THE GERMINATION
AND ROOT ELONGATION OF SELECTED SEEDLINGS.
Western Michigan University, M.A., 1975
Biology
Xerox University Microfilms , Ann Arbor, Michigan 48106
T H IS D IS S E R T A T IO N H A S R EFN M IC R O F IL M E D F X A C T I Y AS R F O F IY /F D
Reprodu ced with permission o f the copyright owner. Further reproduction prohibited without permission.

TABLE OF CONTENTS
CHAPTER
I
II
i n
IV
v
P’ACki
INTRODUCTION
...............................
1
LITERATURE RSVISV;
...........................
3
LETHODS AND MATERIALS
........................
7
Plant Material
............................
7
Sound Source
..............................
7
Growth Conditions
.........................
7
Determination of Germination and Growth
......
9
Statistical Analysis
.......................
9
RES U LT S
....................................
10
Root Elongation
...........................
10
Germination
...............................
11
DISCUSSION
.................
.
...............
37
iii
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

LIST C? TABLr.3
TA3LB
1 lie an length in as of longest root of
germinated cucumbers for control and
sound treated seeds
...........................
13
2 iiiean length in as of longest root of
germinated oats for control and sound
treated seeds
..........................
11
3 mean length in ms of longest root of
germinated wheat for control and sound
treated seeds
................................
15
1 Per cent germination of cucumber for control
and sound treated seeds
.......................
16
5 Per cent germination of oats for control
and sound treated seeds
.......................
17
6 Per cent germination of wheat for control
and sound treated seeds
........................
18
7 Two-way analysis of variance (unbalanced
case) for root elongation data of
cucumbers for runs 1 and 2
.....................
19
8 Two-way analysis of variance (unbalanced
case) for root elongation data of
cucumbers runs 3 and 1
............
9 Two-way analysis of variance (unbalanced
case) for root elongation data of
cucumbers runs 5 and 6
........................
21
1C Two-way analysis of variance (unbalanced
case) for root elongation data of
cucumbers runs 7 and 8
........................
22
11 Two-way analysis of variance (unbalanced
case) for root elongation data of
cucumbers all runs
............................
23
iv
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

TABLE PAGE
12 Two-way analysis of variance (unbalanced
casej for root elongation data of
oats runs 1 and 2
............................
24
13 Two-way analysis of variance (unbalanced
case) for root elongation data of
oats runs 3 and 4
............................
25
14 Two-way analysis of variance (unbalanced
case) for root elongation data of
oats runs 5 and 6
............................
26
15 Two-way analysis of variance (unbalanced
case) for root elongation data of
oats runs 7 and S
............................
27
16 Two-way analysis of variance (unbalanced
case) for root elongation data of
oats all runs
................................
28
17 Two-way analysis of variance (unbalanced
case) for root elongation data of
wheat runs 1 and 2
...........................
29
18 Two-way analysis of variance (unbalanced
case) for root elongation data of
wheat runs 3 ana 4
...........................
30
19 Two-way analysis of variance (unbalanced
case) for root elongation data of
wheat runs 5 and 6
...........................
31
20 Two-way analysis of variance (unbalanced
case) for root elongation data of
wheat runs 7 and 8
.............
32
21 Two-way analysis of variance (unbalanced
case) for root elongation data of
wheat all runs
...............................
33
22 Two-way analysis of variance of cucumber
germination data
.............................
34
23 Two-way analysis of variance of oat
germination data
..........
35
24 Two-way analysis of variance of wheat
germination data
.............................
36
v
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

THE EFFECT OF AUDIBLE SOUND ON THE GERMINATION AND
•ROOT ELONGATION OF SELECTED SEEDLINGS
Introduction
Sound is energy which travels in the fora of sound waves. It is
present throughout the environment so that all living organisms are
exposed to it. lian can detect sounds ranging in intensity from 10“^
watts/m^ to 1 watt/m^. The lower intensity is barely audible while
the higher intensity is pain producing. The range of frequencies
which man can detect is referred to as the audible sound range. Those
sounds of frequencies above the audible range are called ultrasounds.
The frequency of sound is measured in Hertz or cycles per second
and the intensity of sound is expressed in decibels (db). The decibel
is defined according to a logarthmic scale. The intensity level in
decibels r 10 log l/l0 where ID is the intensity of the faintest
audible sound which is 1 0 " ^ watts/m^. This being the case, the
intensity level of the least audible sound is 10 log (10_*^/lC~-^)
which is zero.
kost experiments reported in the literature involve the use of
ultrasound (sound of higher frequency then audible sound). Ultra
sound has very destructive effects on living cells. Yeast and bacterial
cells present in milk have been destroyed upon exposure to ultrasonic
waves (Beckwith and 7/eaver, 1936). The destructive effects of ultra
sound are largely due to cavitation. Cavitation involves the formation
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

of partial vacuums -within the cell due to the separation of intra
cellular structures (Clark and Hill, 1969). Ultrasonic waves are
frequently used in the destruction of microbial cells and the
separation of intra-cellular particles (El'Piner, 1964).
Experiments with audible sound reported in the literature
generally involved animals. Disruption of animal cells by audible
sound may be produced by the explosion of the organism as a result
of the internal release of dissolved gases, or by actual tearing
apart of tissues as a result of rapid alterations of tension and
compression produced in the surrounding medium by the vibrations
(Chambers and Gaines, 1932). Plant experimentation with audible
sound generally concerns effects on germination and inhibition
or stimulation of plant growth.
This work was initiated to examine the effects of audible
sound on the germination and growth of cucumbers, oats, and wheat.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

LITERATURE REVIEW
The effects of audible sound on animals and animal cells has
been investigated in several ways. Chambers and Gaines (1932) found
that the water flea, Daphnia pulex, when exposed to sonic irradiation
in a shell vial filled with water, was killed within one second. The
exoskeleton was shattered, the soft body parts were churned into an
amorphous pulp, and clouds of colloidal material were released into
the surrounding water. 3rys’nenskii and Meskhaeleva (1969) found that
animals subjected to the prolonged action of sound exhibited a lower
coagulation activity of the blood. The sound weakens the defense
reactions which develope after the injection of thrombin. The death
rate amoung the experimental animals was 72.5% while the death rate
for the control animals was 40.9*. Aleksandrovskaya and Chezhenkova
(1970) found that a weak sound (200 c.p.s.) in the rabbit cerebral
cortex enhanced the slow waves and spindles, particularly in the motor
area. In the deep layers of the motor area, the weak sound brought
about an increase in astrocytes. In the deep layers of the projection
area (auditory cortex) the astrocytes decreased in number.
Xoitchev (1969) found that there was an increase of sodium in
the organ of Corti after exposure to sound. This was attributed to
changes in the permeability of excited membranes. Chambers and Harvey
(1931) found that tadpoles of 3ufo punctata when held in shell vials
filled with water, above the vibrator, were killed within one minute.
After one minute of sound treatment, bubbles of gas could be seen in
3
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

the abdominal cavity, and after two minutes the tadpoles actually
exploded. Small fish were killed in three to four minutes and
immature frogs were killed in about ten minutes.
Most of the work on the effects of audible sound on plants
and plant cells concerns growth and germination. Northen and
hacVicar (1939) showed that sound lowered the elasticity of the
cytoplasm of Spirogyra by 20 per cent to 88 per cent. This could
be seen by the displacement of chloroplasts in sound treated
Spirogyra filaments. Singh (1959) reported that the water plant
Hydrilla verticillata had an increased rate of photosynthesis
after exposure to sound waves from an electric bell. 3oth the rate
and the total volume of oxygen evolved from sound-excited Hydrilla
plants were 60 per cent to 100 per cent higher then the control
plants. Gnanam (1959) observed an increase in the photosynthetic
rate of Spirogyra when exposed to sound. The volume of oxygen
evolved in excited plants vras much higher then the control,
showing thereby, that in sound excited plants synthesis and
production of food was higher.
Some rather non-scientific literature has been concerned
with the effect of music on plants. RetaHack (1973) found that
acid rock, after being played to plants for five days, had very
devastating effects on plant growth. She observed a bending of the
plants away from the speaker and a decrease in root developement.
liusic played by Ravi Shankar had no effect on root developement
and the plants were found to be bending toward the speaker. Some
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

5
reference to the effects of music on plants has been recorded in
scientific literature. Ponniah (1958) played a single note to
Mimosa pudica, Impatiens balsamina, Tagetes erecta, and Hydrilla
verticillataa She found an increase in root developement, total
number of leaves, and length of branches. Singh and Ponniah (1955)
found that the musical sound of the Veena stimulated the growth
of balsam plants in all parameters measured.
Lisenkov (1966) found that seeds of Siberian larch exposed
to sound treated water had an increase in ground germination,
increased frost resistance, and better growth. The enzyme activity
of the germinated seeds was also increased under certain conditions.
Amylase activity increased when the seeds were soaked in sound
treated water for periods between 0.5 and 1.0 hours.
Weinberger and Measures (1968) exposed spring and winter
wheats (Triticum aestivum) to a single audible frequency of
sound at a given time. They found better germination and an
increase in growth in winter wheat (var. Rideau) and in some
cases this was temperature or frequency dependent. Spring wheat
(var. Marquis) showed an increase in germination at 2 degrees C
and 10 degrees C, but not at 25 degrees C. The growth of spring
wheat was found to vary with different treatments. It was suggested
that the increase in germination at 2 degrees C and 10 degrees C
may involve the production of ethylene.
The same workers in another experiment using spring wheat
found a significant increase in some of the parameters measured
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

when 3^0 Hz and 5,000 Hz were used. However, plant growth was not
significantly stimulated by treatment with either 1,250 Hz or
12.000 Hz sound frequencies (keasures and Weinberger, 1970)•
Some negative effects of sound on growth have also been
reported in scientific literature. Weinberger and Das (1972)
found that continuous exposure of Scenedesmus obtusiuscuius to
1.000 Hz resulted in a decrease in the rate of cell division. A
decline of 15 per cent was obtained in 18 hours. The normal rate
of cell division did not return until two life cycles had elapsed.
The Scenedesmus cells were most sensitive to sonic shock in the
early part of their life cycle. Yfoodlief et al., (1969) also found
some negative effects of sound on plants. Tobacco plants subjected
to random noise showed a decline in growth rate of over 10 per cent.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

METHODS AND MATERIALS
Plant Material
The three types of certified seeds used in these experiments,
wheat (Triticua aestivum), oats (Avena sativa var. AuSable), and
cucumber (Cucumis sativis var. marketer) were obtained from Farm
Bureau Services Inc., Kalamazoo, Michigan. VJhen not being used for
experimentation all seeds were stored in a refrigerator at approx
imately 8 degrees C.
Sound Source
Sound at a frequency of 600 Hz was generated by an audio-
oscillator^- connected to an amplifier^. Sound intensity was
measured by using a sound level meter3 using the 'G' weight. The
audio-oscillator was adjusted to give sound of the intensity of
102 plus or minus 3 decibels in each of the two growth chambers.
In all experiments a nine inch speaker was used. The speaker was
suspended by heavy strings near the center of the growth chamber.
Growth Conditions
Both control series and experimental series consisted of
^-Audio-oscillator made by Hewlett Packard Co., Pala Alto,
California (Model 200 CD).
^Amplifier made by the David Bogen Co., New York, New York
(jiodex m l j.
3Sound level meter made by the General Radio Co., Nest
Concord, Massachusetts.
7
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

8
5 plastic dishes-*- (155 on by 63 mm). Each dish was lined with Armstrong
ft6 filter paper^ and was moistened with 15 ml of distilled water. A
total of 20 seeds were randomly scattered in each dish giving 100
seeds for each run. The plastic containers were then placed in clear
polyethylene bags3 which were closed by using wire twists. After ail
dishes were moistened and planted, they were placed in Sherer Controlled
Environment Chambers^-. Experiments in which cucumbers were tested and
those in which oats were tested were run simultaneously. The experiments
with wheat were run separately since there was not enough room in
the growth chamber to run 3 sets of dishes at the same time.
The dishes were placed near the center of the growth chamber
in a circular pattern. The growth chambers (experimental and control)
were kept in constant darkness at a temperature of 20-21 degrees C.
The experimental series was continuously exposed to sound. The control
series was ran in a second growth chamber under exactly the same
conditions and at the same time. The only sound in the control
chamber was due to normal background noise. After 5 days all seeds
■were removed and measured. A total of 8 separate runs, 4 replications
^Plastic containers were obtained from 3radley Industries,
Inc., Franklin Park, Illinois.
2?aper manufactured by the Armstrong Cork Co., Lancaster,
Pennsylvania.
^3ags made by Union Carbide Corp., Consumer Products Division,
270 Park Avenue, New York, New York, and sold under the trade name
of 'Glad*.
^Sherer-Gillett Co., Marshall, Michigan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

of the experiment, were made for each of the 3 types of seeds.
The growth chambers were interchanged for each succeeding run
sc that the 1 experimental' chamber became the 'control1 chamber
on the next run.
Determination of Germination and Growth
Any seed was considered to have germinated if its root reached
a length of 1 mm or longer. Growth was determined by using the
method of Thompson et al., (1915) where the longest root of each
germinated seed was measured in mm. A mean root length was determined
for both control and experimental groups by using only root lengths
of seeds that did germinate.
Statistical Analysis
Data on root elongation were analyzed by means of library,
program # 1.9.2. using Western Michigan Universities PDP-10
computer. This program is a two-way analysis of variance (unbalanced
case) where factor 1 is the difference between control and experimental
treatments and factor 2 is the .difference between rims. Seeds that
did not germinate were not included in the analysis. Data on
germination were analyzed using library program# 1.9.1. (version 2).
This program is also a two-way analysis of variance. It shows the
significance of control/experimental, replication, and growth chamber
differences. It does not consider within cell variability. In both
programs used a probability level was calculated from the 'F' value.
Reproduced with permission of the copyright owner. Further reproduction prohibited w ithout permission.

RE SU LTS
Root Elongation
Tables 1, 2, and pages 13, 14, and 15 show the average length
of the longest root of germinated cucumber, oat, and wheat seeds under
both control and experimental conditions. In the case of cucumbers and
oats the grand mean (GM) of the control group is significantly greater
then the grand mean of the experimental group. In the case of wheat the
grand mean of the control group is not significantly greater then the
grand mean of the experimental group. Tables 7-21 on pages 19-33 show
the results of statistical analysis.
Cucumber seeds
YVith cucumbers (see Table 1, page 13), the grand mean of the
control group is 56.61 mm while the grand mean of the experimental
group is 47.09 The difference observed is significant at the 0.1
per cent level (see Table 11, page 23). A significant difference (at
the 3.5 per cent level) is found between the 8 runs of the experiment.
Interaction (which determines if differences are due to experimental
conditions or some other factors) is significant at above the 23 per
cent level. Since interaction is not significant at below the 5 per
cent level, no further analysis of root elongation data for cucumbers
is needed.
Oat seeds
The data for oats (see Table 2, page 14) is similar to that
for cucumber showing an inhibition of growth in the sound treated
10
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

11
•group. The grand mean of the control group is 112.17 mm and the grand
mean of the experimental- group is 99.23 am. The difference between
the two is a significant difference at below the 0.1 per cent level
(Table 16, page 28). There is a significant difference between the
8 runs (below the 0.1 per cent level). The interaction is not
significant being above the 70 per cent level. Once again, since
interaction is not significant no further analysis of root elongation
data for oats is needed.
Wheat seeds
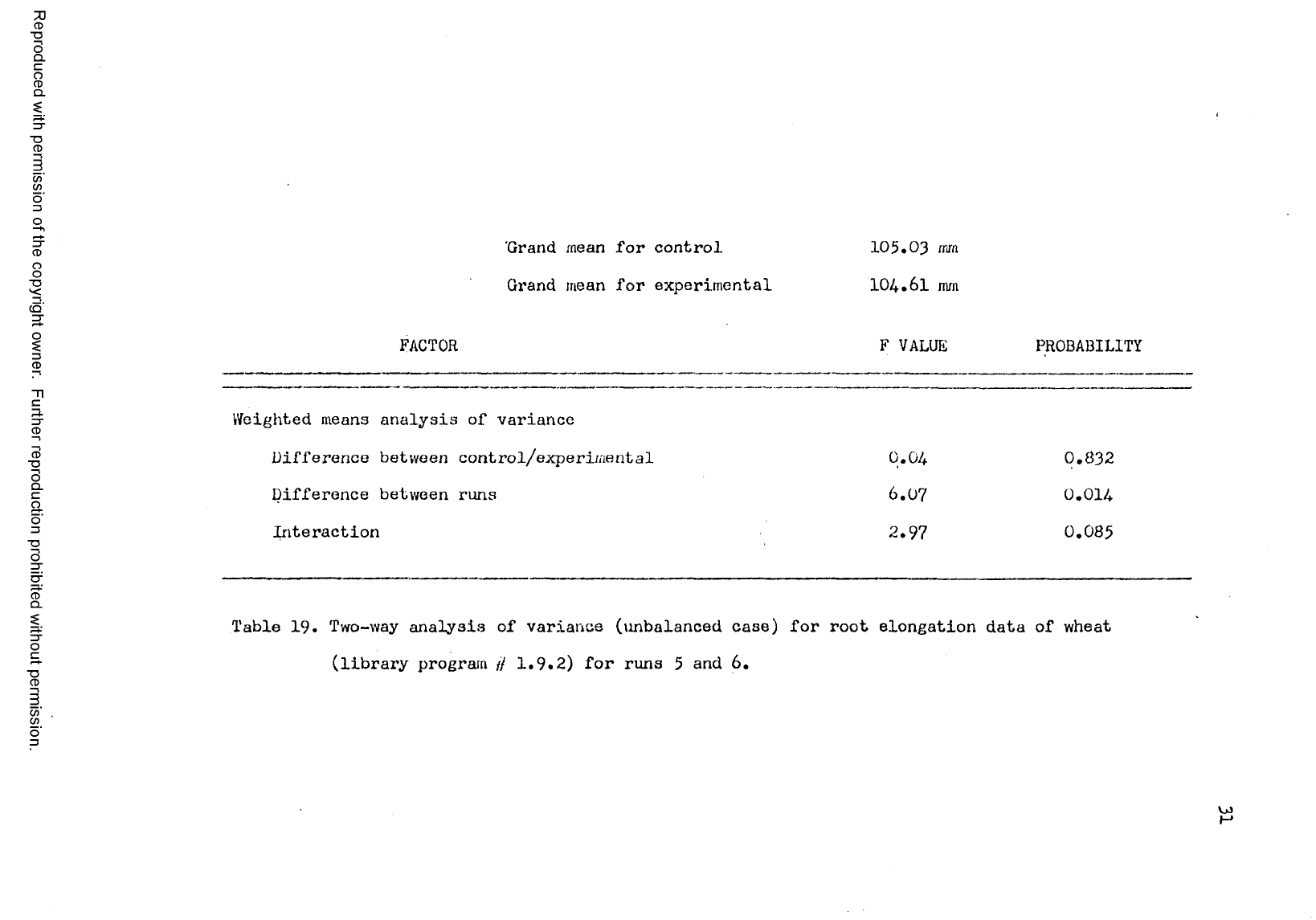
The data for wheat (see Table 3, page 15) are different than
the data for cucumbers and oats showing no real inhibition of growth
in the sound treated group. The grand mean of the control group is
105.03 mm and the grand mean of the experimental group is 101.61 mm.
The difference between the two is not significant (Table 19, page 31).
There is a significant difference between the 8 runs (below the 1.1
per cent level). The interaction is significant only above the 8.5
per cent level. Since the difference between the control and sound
treated -wheat seeds is not significant no further analysis of the
data is required.
Germination
Tables 1, 5, and 6; pages 16, 17, and 18 show the per cent
germination for cucumbers, oats, and wheat. Tables 22, 23, and 21;
pages 31, 35, and 36 show the level of significance for-the differences.
In cucumbers, oats, and wheat there is no significant difference
in per cent germination for either growth chambers, replications,
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

12
or control/experimental differences.
Cucumber seeds
The data indicate that there is no significant difference in
per cent germination between control and experimental plants (22.1
per cent level), between replications (36.5 per cent level), or
between growth chambers A and B (22.1 per cent level).
Oat seeds
The data indicate that there is no significant difference in
per cent germination between control and experimental plants (84.4
per cent level), between replications (8.0 per cent level), or
between growth chambers A and B (55.5 per cent level).
VTheat seeds
As in cucumbers and oats, the data indicate that there is no
significant difference in per cent germination between control and
experimental plants (18.5 per cent level), between replications
(18.5 per cent level), or between growth chambers A and 3 (42.5
per cent level).
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Replications
G il
56.24
56.61 62.96
59.08
47.95
Control
53.81
51.85
52.11
38.31
Sound
Runs
Table 1. iiean length in rnrn of longest root of germinated cucumbers for control
and sound treated seeds. GM = Grand Mean. (Significant to 0.1 mm).
H*
VO

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Control
Sound
Table 2.
Replications
GM 1
2
3
4
112.17 95.47
119.94
109.51
104.14
107.78
121.53
122.85
116.11
99.23
83.97 104-93
98.56
86.78
93.74 108.97
108.95
107.93
1 2
3
4
5
6
7
8
Runs
Mean length in mm of longest root of germinated oats for control and
sound treated seeds. GM = Grand Mean. (Significant to 0.1 mm).
£

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Replications
GM
1
2
3
4
Control
105.03
107.04 107.76
106.15
100.62
100.85
102.79
108.78
106.26
Sound
104.61
105.53 109.33
97.35
107.90
96.88
107.86
102.52 109.52
1
2
3
4
.......
5
6
7
8
Runs
Table 3« Mean length in ram of longest root of germinated wheat for control and
sound treated seeds. GM = Grand Mean. (Significant to 0.1 nun)

16
to
c
o
•H
P
O
•H
r-H
a
©
os
-O'
CO
O'
-a-
O'
CO
'O
O'
86
c-
MO
cc
O
O'
O'
CO
CO
o
CO
O'
o
1—1
-<}■
!>-
O'
O'
02
o 'O
i
O' O'
CO
CO
o 02
O'
O'
rH
O'
O'
O'
H
o
o
r-i
CO
-a
• •
33
ir\
UO
O'
O'
,
o
u
•c
-p c
c
3
o
o
o
CO
to
a:
•
to
T3
©
©
to
•o
o
-p
CO
to
u
p
•u
o
3
0
to
•o
c
to
H
o
u
p
3
o
o
u
o
a
3
CD
XI
s
3
o
3
o
a
o
c
o
P
p
to
•
3
3
•p
©
a
©
5
tD
CO
"O
3
p ©
3
©
33
o
11
©
a
33
•
-*
a>
r—l
XJ
©
E-*
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Cl
C
o
-P
CO
o
r—<
a
a>
ox
S t
i a
CO
0 6
CO
O i
O '
c a
O '
c -
tr
c -
'O
e e
co
s t
C\i
O '
O '
IA
o O '
- t
O '
CO
c\j
i
1
i
CO 00
CA
CO
00
O '
C\J CM
CO
O '
iH
c -
r_
IA
0 0 00
o
o
o
!A
•
•
O O '
O '
CO
CO
o
T3
- p
c
3
o
o
o
CO
03
I
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Table 5. Per cent gemination of oats for control and sound treated seeds.

1 8
03
C
o
•H
-P
ca
o
•r!
r-i
a
0
1
- ' t
9 5
1 0 0
CO
1 0 0
9 7
C '-
CO
O ' O '
r A
C '-
o
I A
o
o
rH
CO
-4 -
O '
O '
CM
i r \
O '
C A
O '
O '
CO CM
O ' O '
rA
t>
i a
O '
O '
i—i
a
o
o
vO
•
•
3
c -
O '
o
r—I
o
u
TJ
p
d
c
3
o
r o
O
r n
CO
as
03
"O
a)
©
03
TO
0)
P
£3
a)
u
p
•a
c
3
o
03
•o
C
CO
H
o
P
c
o
u
JC.
o
CH
P
CO
©
x :
s
o
c
o
■H
P
CO
•
c
C
•H
CO
d
0
s
©
GO
c
P
ca
c u
©
O
o
11
s*
©
a. 3
•
'O
0
rH
jQ
G3
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control
Grand mean for experimental
56.61 mm
47*09 nun
FACTOR
F VALUE
PROBABILITY
Weighted means analysis of variance
Difference between control/experimental
20.74
0.000
Difference between runs
44.28
0.000
Interaction
1.73
0.190
Table 7. Two-way analysis of variance (unbalanced case)
for root elongation data of cucumbers
(library program jj 1.9.2) for runs 1 and 2.
M
vO

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control
Grand mean for experimental
56.61 mm
47.09 mm
FACTOR
F VALUE
PROBABILITY
Weighted means analysis of variance
Pifference between control/experimental
48.52
0.000
Difference between runs
7,88
0.005
Interaction
1.20
0.275
Table 8 . Two-way analysis of variance (unbalanced case) for root elongation data of cucumbers
(library program // 1 .9 *2) for runs 3 and 4 .
8

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control
Grand mean for experimental
56.61 nan
47.09 mm
FACTOR
F VALUE P R O B A B IL IT Y
Weighted means analysis of variance
Difference between control/experimental
11.92
0.001
Difference between runs
16.18
0.000
Interaction
2.70
0.101
Tabl e 9. T w o - w a y a nalysis of varian ce (u n b a l a n c e d case) fo r root e l o n g a t i o n data of c ucum be rs
(libr ary p r o g r a m // 1.9.2) f o r runs 5 and 6.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control
Grand wean for experimental
5o.6i, mm
47.09 hum
FACTOR F VALUE
PROBABILITY
Weighted means analysis of variance
Difference between control/experimental
13.51
0.000
Difference between rims
1.72
0.191
interaction
0.18
0.668
Table 10. Two-.vay analysis of variance (unbalanced case) for root elongation data of cucumbers
(library program y 1 ,9.2 ) for runs 7 and 8 .
Jo

Reproduced with permission of the copyright owner. Further reproduction prohibited without perm ission.
Grand mean for control.
Grand mean for experimental
56.61 mm
47.09 mm
FACTOR F VALUE
PROBABILITY
Weighted means analysis of variance
Difference between control/experimental
Difference between runs
Interaction
36.66
0.000
A.75
0.035
1.47
0.232
Table 11. Two-way analysis of variance (unbalanced case) for root elongation data of cucumbers
(library program ft 1 .9 .2 ) all runs.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control 112.17 jnm
Grand mean for experimental 99*23 inm
FACTOR F VALUE PROBABILITY
Weighted means analysis of variance
Difference between control/experimental
•15.41
0.000
Difference between runs
45*27
0.000
Interaction
0.27
0.604
Table 12. Two-way analysis of variance (unbalanced case) for root elongation data of oats
(library program if 1 .9*2 ) for runs 1 and 2 .
£

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control
Grand mean for experimental
112.17 mm
99.23 mm
FACTOR
F VALUE PROBABILITY
Weighted means analysis of variance
Difference between control/experimental
13.19
0.000
Difference between runs
4.83
0.029
Interaction
0.68
0 . 4 H
Table 13. Two-way analysis of variance (unbalanced case) for root elongation data of oats
(library program # 1 .9 .2 ) for runs 3 and 4 .
M

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control
112,17 mm
Grand mean for experimental
1
99*23 mm
FACTOR F VALUE
PROBABILITY
Weighted means analysis of variance
Difference between control/experimental
17.91
0.000
Difference between runs
21.25
0.000
Interaction
0.05 0.815
Table 1^. Two-way analysis of variance (unbalanced case)
for root elongation data
of oats
(library program ft 1 .9 .2 ) for runs 5 and 6 .
fo
O '

Grand mean for control
Grand mean for experimental
112.17 mm
99.23 mm
FACTOR F VALUE P R O B A B IL IT Y
Weighted means analysis of variance
Difference between control/experimental
9.74
0.002
Difference between runs
1.20
0.274
Interaction 0.66
0.419
Ta b l e 15. T w o - w a y a nalys i s of v ari a n c e ( u nb al a n c e d case) f o r r o ot e l o n g a t i o n d a ta of oats
(l i brar y p r o g r a m
it
1. 9. 2) f o r runs 7 a n d B.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control 112.17 mm
Grand mean for experimental 99*23 mm
FACTOR F VALUE PROBABILITY
Weighted means analysis of variance
Difference between control/experimental
32.05
0.000
Difference between runs
21.82
0.000
Interaction 0.15
0.702
T a b l e 16. Tw o - w a y an a l y s i s of v a r i a n c e ( u nb a l a nce d case) f or root e l on g a t i on d a ta of oats
(library p r o g r a m
if
1.9.2) all runs.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control
Grand mean for experimental
105o03 mm
104.61 mm
FACTOR
F VALUE
P R O B A B IL IT Y
Weighted means analysis of variance
Difference between control/experimental
Difference between runs
Interaction
0.00
0.60
0.28
0 . 9 9 3
0.440
0.598
Table 17. Two-way analysis of variance (unbalanced case) for root elongation data of wheat
(library program // 1 .9 .2) for runs 1 and 2 .
M
v O

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control
Grand mean for experimental
FACTOR
105.03 mm
104.61 mm
•F VALUE
PROBABILITY
Weighted means analysis of variance
Difference between control/experimental
0.09
0.767
Difference between runs
0.97
0.326
Interaction
9.92
0.002
Table 18. Two-way analysis of variance (unbalanced case)
for root elongation data
of wheat
(library program rf 1.9.2) for run3 3 and 4.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
'Grand mean for control 105.03 mm
Grand mean for experimental 104.61 mm
FACTOR F VALUE PROBABILITY
Weighted mean3 analysis of variance
Difference between control/experimental
0.04
0.832
Difference between runs
6.07 0.014
Interaction
2.97
0.085
Table 19. Two-way analysis of variance (imbalanced case) for root elongation data of wheat
(library program // 1.9.2) for runs 5 and 6.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control
Grand mean for experimental
10^.03 mm
lOA.bl mm
FACTOR
F VALUE
PROBABILITY
Weighted means analysis of variance
Difference between control/experimental
0.31
0.579
Difference between runs
O .69
0.408
Interaction 3.09 0.079
T a b l e 20. T w o - w a y analys is of v a r i a n c e (unb a lan c ed case) for root e l o n g a t i o n d a t a of wheat
( l ib r a r y p r o g r a m # 1.9.2) fo r runs 7 a n d 8.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Grand mean for control
105.03 mm
Grand mean for experimental
10A.61 mm
FACTOR
F VALUE
PROBABILITY
Weighted means analysis of variance
Difference between control/experimental
0.17 0.683
Difference between runs 1 0.38
0.541
Interaction
0.00
1.000
Table 21. Two-way analysis of variance (unbalanced case)
for root elongation data
of wheat
(library program # 1.9.2) all runs.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
f a c t o r f v a l u e p r o b a b i l i t y
Cucumber
Difference between control/experimental
1.53
0.221
Difference
between replications
1.18
0.365
Difference
between chambers A and B
1.53
0.221
Table 22. Two-way analysis of variance of cucumber germination data (library program # 1.9*1
(version 2) ).
•t--
i

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
FACTOR F VALUE PROBABILITY
Oats
Difference
between control/experimental
0.0/+
0.844
Difference between replications
3.17
0.080
Difference between chambers A and B
0.35 0.555
Table 23. Two-way analysis of variance of oat germination data (library program # 1.9.1
(version 2) ).
vo
vn

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
FACTOR F VALUE PROBABILITY
Wheat
Difference between control/experimental
1.79 0.185
Difference between replications
1.79
0.185
Differences between chambers A and B
0.65 O.A25
Table 2k* Two-way analysis of variance of wheat germination data (library program // 1.9.1 (version
2) ).
VjJ
O '

DISCUSSION
Cucumbers and oats treated with 600 Hz audible sound showed a
significant inhibition of root elongation. Wheat did not show any
significant inhibition of root elongation. No significant change in
per cent germination was observed for any of the experimental plants.
In all runs, considerable variation is observed in both control
and experimental groups. In cucumbers, the control plants show a low
mean of 47.95 mm and a high mean of 62.96 mm: experimental plants
show a low mean of 38.31 mm and a high mean of 53*81 mm. In oats,
the control plants show a low mean of 95.47 mm and a high mean of
122.85 mm. The experimental plants show a low mean of 83.9? mm and a
high mean of 108.97 mm. Wheat control plants show a low mean of 100.62
mm and a high mean of 108.78 mm. Experimental wheat plants show a
low mean of 96.88 mm and a high mean of 109.52 mm.
Since these variations cannot be attributed to growth chamber
differences (see Tables 22, 23, and 24; pages 34* 35* and 36), some
other factors must be involved. It seems logical that morphological
variation in seeds may be a factor. Some selection in seeds was done
during the course of the experiment. For example, all seeds that
were obviously much larger or smaller then the 'average1 seeds were
not used in the experiment. Any broken or rotten seeds were discarded.
However, considerable variation in runs still occurred. Perhaps the
answer to this problem would be some type of screening apparatus. A
series of two screens could be placed one below the other so that
37
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

very large seeds would not pass through the top screen. The seeds
collected on the bottom screen (which would have smaller openings
then the top screen) would be about the sane size with the very
snail seeds falling through both the first and second screens. It
also seems possible that size variations for runs would be smaller
if the seeds were allowed to grow for a longer period of time. It
was observed in other experimentation with Cyprus esculentus
(nutsedge) that tubers which were heavier and larger in size were
considerably larger then the lighter smaller tubers a few days
after germination. However, after several weeks of growth it was
observed that the differences in size were considerably less noticeabl
At 600 Hz of sound, treated cucumber and oat seeds showed a
significant inhibition of root elongation 'while wheat seeds did not.
Work previously done in this lab (Ellis, 1973) at 300 Hz sound
showed an inhibition of root elongation in wheat. However, it could
not be stated conclusively that the inhibition was due to sound
since interaction was significant. Experiments now in progress
(Gyimah) indicate that there is no significant inhibition of root
elongation in wheat, oats, or cucumber at 900 Hz or at 1,200 Hz.
Weinberger and Measures (1970) found an increase in growth in Marquis
spring -wheat at 300 Hz of sound. They found no effect on growth at
1,250 Hz of sound treatment. The differences observed in the growth
responses of different species of plants at different frequencies
of sound treatment is not disturbing. There are several possible
explanations for the occurrence of these varied responses.
In order for sound to effect growth, it seems logical that the
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

39
sound waves must penetrate the seeds. Possibly the nake up of the seed
coat varies with different species. A thick seed coat would make it sore
difficult for sound waves to enter the seed then a thin seed coat. It
would be of interest to pursue this in further experimentation. It is
also possible that there are different biochemical pathways in different
species of plants. Some of these pathways may be disrupted by sound
waves while others may be unaffected. It right be that an enzyme or
an intermediate metabolic product could be broken down which would
result in the destruction of a particular pathway necessary for normal
growth to occur. Within the same plant species, it is quite possible
that only a specific frequency of sound would have the ability to
disrupt a particular metabolic pathway. It ’would be useful to carry
on further experimentation in which the amino acid levels of sound
treated and control seeds could be measured to see if there are any
significant differences.
Another interesting question is how do sound waves inhibit growth
in plants when this is seen to occur. Many investigators have attributed
the destructive effects of sound waves on cells to cavitation (Chambers
and Gaines, 1932). Cavitation involves the tearing apart of a column
of fluid, and the production in it of regions of vapor or gases. The
effects of cavitation are much more evident at very high frequencies
of sound (ultrasound) then at audible frequencies.
Why sound waves do inhibit growth in some species, but may have
no effect or may even stimulate grov/th in other species of plants is
not really known. Weinberger and Measures (1970) found differences
in growth in the same species of wheat when the frequency of sound
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

40
was varied. Below a certain frequency no inhibition of growth or
germination could be seen. Above this frequency inhibition of growth
and germination did occur. The frequency and amount of energy required
to produce these effects appears to vary widely between species.
In this experiment there is no significant interaction (Tables
11, 16, and 21; pages 23, 28, and 33). This indicates very strongly
that the observed effects, i.e. inhibition of root elongation in
cucumbers and oats, is due to sound, i.'o inhibition of root elongation
is observed for wheat. The data for cucumbers and oats, however, point
to a most dramatic inhibition of root elongation.
Germination in cucumbers, oats, ana wheat treated by 6G0 Hz sound
is not significantly effected (Tables 22, 23, and 24; pages 34, 35,
and 36). In cucumbers the data is significant at the 22 per cent level,
in oats at the 84 per cent level, and in wheat at the 18 per cent
level.
In conclusion, I have rejected the null hypothesis — Uc = Ue
(no significant difference between control and experimental runs
exists) for root elongation in cucumbers and oats. On the basis of
a statistical analysis which includes all runs, I am able to state
conclusively that the reduction in root elongation is due to sound
treatment. However, the null hypothesis cannot be rejected for wheat
since no significant inhibition of root elongation occurred. In future
experiments it would be of value to be more selective in the choice
of seeds. If only those seeds which were morphologically and genetically
similar were used (such as screened seeds; there might be less
variability between runs. Experiments should be designed which
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

41
consider the changes (if any) in anino acid concentration between
control and sound treated seeds. A microscopic examination of seed
coats to determine differences in thickness would be of value. Finally,
it would be of interest to run either cucumbers and wheat or oats and
wheat at the same time. In doing this, a species which did show root
inhibition and one which did not show root inhibition could be run
simultaneously. The null hypothesis for the effect of 600 Hz sound
on germination — U c = Ue— is not rejected.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

l it e r a t u r e c i t e d
Aleksandrovskaya, M. M. and R. A. Chezhenkova. 1970.
Electrofiziologicheskie i morfologicheskie issledovaniya
raznykh oblastei kory bol'shikh polusharii vozdeistvii
slabym zvukom. (Electrophysiological and morphological
study of different cerebral cortical parts during weak
sound stimulation). Fiziol. Zh. SSSR. Im. I, K. Sechenova.
5o(3):312-317.
Chambers, L. A. and Gaines N. 1932. Some effects of intense
audible sound on living organisms and cells. Jour, of
Cellular and Comparative Physiology. 1:431-469.
Ellis, George Feme. 1973* The Effect of Audible Sound on the
Germination and Root Elongation of Oats and Wheat. Master’s
Thesis.
El'Piner, I. Y. 1970. Nekavitatsionnyi ul'trazvuk i ego biologicneskoe
deistvie. (Non-Cavitational ultrasound and its biological
effect). 3io Fizika 15(3):333-343.
Frings, H., C. H. Allen, and I. Rudnick. 1948. The physical
effects of high intensity air-borne ultrasonic waves on
animals. Jour, of Cellular and Comparative Physiology.
31:339-358.
Gnanam, A. 1959. Activation of photosynthesis in Spirogyra by
sound waves of electric bell. Prod. Symp. Algae (New Delhi).
144-146.
Goldman, D. E. and V7. W. Lepeschkin. 1952. Injury to living cells
in standing sound waves. Jour, of Cellular and Comparative
Physiology. 40:255-26?.
Goldman, D. E. and 11. '3. Lepeschkin. 1957. Injury and recovery of
Spirogyra exposed to ultrasound. Experimental Cell Research.
12:507-517.
Koichev, K. A. 1969. Elektronnomikroskopicheskoe issledovania
raspredeleniya natriya v kortievom orange zhivotnykh, nakhodyash
chikhsys v sostoyanii otnositel'nago pokoya i v usloviyakh
zvukovogo voxdeistviya. (Electron microscope study of sodium
distribution in the organ of Corti of animals under conditions
of relative rest and after exposure to sound)• Tsitologiya
11(5):537-541.
Lepeschkin, Vi. It. and D. E. Goldman. 1952. Effects of ultrasound on
cell structure. Jour, of Cellular and Comparative Physiology.
40:383-397.
42
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

43
Lisenkov, A. F. 1966. Vliyanie ozvuchennoi vody na seaena drevensnykh
rastenii. (The effect of sound-treated water on seeds of woody
plants). Soviet Plant Physiology. 13(4):728-729.
Measures, M. and P. Weinberger. 1970. The effect of four audible
sound frequencies on the growth of Marquis spring wheat. Canadian
Jour, of Botany. 48:659-662.
Northen, H. T. and H. Mac Vicar. 1939. Studies of protoplasmic
structure in Spirogyra. VI. Effects of sound and electricity
on elasticity. Cytologia. 10(1/2):18-21.
Ponniah, 3. 1958. On the effect of musical sound of stringed instruments
on the growth of slants. Indian Sci. Congr. Assoc. Proc. 42:
255.
Retallack, Dorothy. 1973- The sound of music and plants. Santa Monica,
De Vorss and Co., 93 p.
Singh, T. C. N. 1959. On the activation of photosynthesis in Kydrilla
verticullata Presl. by sound waves of an electric bell. Proc.
IX International 3ot. Congr., Montreal. Aug.
Singh, T. C. K. and S. Ponniah. 1954. On the effect of musical sound
of the violin on the growth of Mimosa puaica L. Proc. Indian
Sci. Congr. 161.
Singh, T. C. N. and S. Ponniah. 1955. Effect of musical sound of
veena on balsam plants. Proc. Bihar. Acad. Agril. Sci. 4:122-
125.
Singh, T. C. N. and S. Ponniah. 1955a. On the response of the structure
of the leaves of balsam and mimosa to the musical sound of
violin. Proc. Indian Sci. Congr. Assoc. 42(3):254.
Weinberger, P. and S. Das. 1972. The effect of audible and low
ultrasound frequency on the growth of synchronized cultures of
Scenedesmus obtusiuscuius. Can. Jour, of Botany. 50:361-365.
Weinberger, P. and M. Measures. 1968. The effect of two audible
sound frequencies on the germination and growth of a spring
and winter wheat. Can. Jour, of Botany. 46:1151-1162.
Woodlief, C. B., R. H. Royster, and 3. K. Huang. 1969. Effect of
random noise on plant growth. Jour. Acous. Soc. of America.
46(2 part 2):481-482.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
